inst/doc/wrightscape_github.md
In cboettig/wrightscape: wrightscape
% Detecting a release of constraint in Labrid fish
% Carl Boettiger, Jeremy Beaulieu and Peter C. Wainwright
%
Introduction
The release of constraint hypothesis
Key innovations in parrotfish
(Price et al. 2010)

Models for a release of constraint
The Ornstein-Uhlenbeck (OU) process is a stochastic, mean-reverting
process commonly used in the comparative phylogenetics context to model
the evolution of a trait under constraint (Hansen and Martins 1996). The
model has been generalized to the case of multiple optima in (Butler and
King 2004) and recently to the case of differing strengths of selection
(Beaulieu et al. 2012). This latest extension is perhaps most
interesting when used to detect a change in selection strength in a
sub-clade following a potential innovation. This would predict and
increase in the rate disparity increases in some focal traits in the
sub-clade relative to the base rate observed. We will refer to this
model in which the strength of stabilizing selection decreases as the
release of constraint model.
Unfortunately, this basic pattern corresponds to a similar scenario that
does not involve a release of constraint. It is commonly postulated that
a trait’s evolutionary pattern may correspond best to a Brownian motion
(BM) process without no central tendency imposed by the constraint in
the OU model. If the basic Brownian rate parameter increases at the time
of the innovation, a pattern of increased growth in disparity can still
be observed without the corresponding mechanism of a release of
constraint. Modeling a change in a Brownian rate parameter was first
introduced in @O’Meara2006 in the software Brownie; hence we will
refer to this as the Brownie model.
Though the processes are not identical, the exhibit remarkably similar
patterns. Figure 2 illustrates the Brownie and release of constraint
models through 500 replicate simulations of each. In the top panel, BM
and OU processes with comparable parameters are shown for reference. In
both the Brownie and release of constraint models, the same variance has
been reached at the time of the shift and at the time the simulation
ends. The distinguishing feature in the release of constraint model is
similar to the distinction between a BM and OU processes – before the
shift occurs, the trait dynamics have begun to approach an equilibrium
that balances the diversification process against the constraint. This
corresponds to traits values more closely reflecting a match to their
environment than to their evolutionary history. After the shift in
selection occurs, the traits begin to explore outside the range
previously possible under the strong constraint.
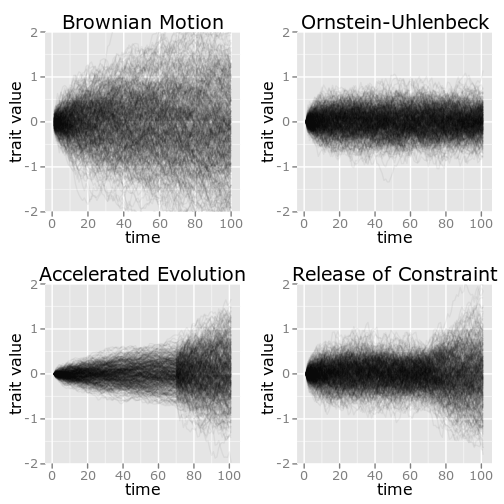
By contrast, the Brownie model shows a sharper transition at this
boundary. A close look at the figure shows a shock front at the moment
of this transition.\
Whereas in the release of constraint model, the trajectories only loose
their central bias, but otherwise continue to make similar step sizes,
in the Brownie model the entire tempo of the evolutionary process has
changed, taking bigger steps in both directions. It is these subtle
differences in the patterns of the evolutionary processes implied by the
different models that we seek to tease apart. To obtain the most
powerful statistical comparison between the two models that accounts for
the uncertainty in the model estimate, we use the method described in
Boettiger, Coop, and Ralph (2012) which uses a bootstrap simulation
approach to compare likelihood ratios of the models.
Results
Estimating the release of constraint model
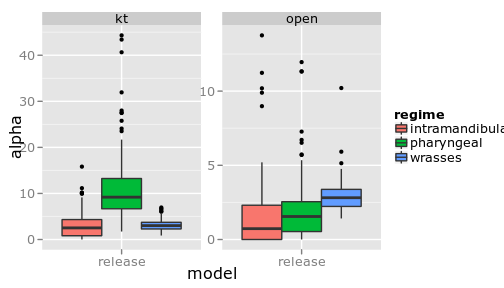
Comparing models

(Show that Brownie model rejects the OUCH model/thetas?)
Discussion
Our analysis identified two functional traits in the labrid jaw
morphology that show clear evidence of a release of constraint.
Functional innovations…
Acknowledgements
This work was supported by a Computational Sciences Graduate Fellowship
from the Department of Energy under grant number DE-FG02-97ER25308 to CB
and NSF grant DEB-1061981 to PCW.
References
Beaulieu, Jeremy M., Dwueng-Chwuan Jhwueng, Carl Boettiger, and Brian C.
O’Meara. 2012. “Modeling Stabilizing Selection: Expanding the
Ornstein-Uhlenbeck Model of Adaptive Evolution.” Evolution (mar).
doi:10.1111/j.1558-5646.2012.01619.x.
http://doi.wiley.com/10.1111/j.1558-5646.2012.01619.x.
Boettiger, Carl, Graham Coop, and Peter Ralph. 2012. “Is your phylogeny
informative? Measuring the power of comparative methods.” Evolution
(jan). doi:10.1111/j.1558-5646.2012.01574.x.
http://doi.wiley.com/10.1111/j.1558-5646.2012.01574.x.
Butler, Marguerite A., and Aaron A. King. 2004. “Phylogenetic
Comparative Analysis: A Modeling Approach for Adaptive Evolution.” The
American Naturalist 164 (dec): 683–695. doi:10.1086/426002.
http://www.jstor.org/stable/10.1086/426002.
Hansen, Thomas F., and E. P. Martins. 1996. “Translating between
microevolutionary process and macroevolutionary patterns: the
correlation structure of interspecific data.” Evolution 50: 1404–1417.
Price, Samantha a, Peter C. Wainwright, David R. Bellwood, Erem
Kazancioglu, David C. Collar, and Thomas J. Near. 2010. “Functional
innovations and morphological diversification in parrotfish.”
Evolution; international journal of organic evolution 64 (oct):
3057–68. doi:10.1111/j.1558-5646.2010.01036.x.
http://www.ncbi.nlm.nih.gov/pubmed/20497217.
cboettig/wrightscape documentation built on May 13, 2019, 2:12 p.m.
R Package Documentation
Browse R Packages
We want your feedback!
Note that we can't provide technical support on individual packages. You should contact the package authors for that.
% Detecting a release of constraint in Labrid fish % Carl Boettiger, Jeremy Beaulieu and Peter C. Wainwright %
Introduction
The release of constraint hypothesis
Key innovations in parrotfish
(Price et al. 2010)
Models for a release of constraint
The Ornstein-Uhlenbeck (OU) process is a stochastic, mean-reverting process commonly used in the comparative phylogenetics context to model the evolution of a trait under constraint (Hansen and Martins 1996). The model has been generalized to the case of multiple optima in (Butler and King 2004) and recently to the case of differing strengths of selection (Beaulieu et al. 2012). This latest extension is perhaps most interesting when used to detect a change in selection strength in a sub-clade following a potential innovation. This would predict and increase in the rate disparity increases in some focal traits in the sub-clade relative to the base rate observed. We will refer to this model in which the strength of stabilizing selection decreases as the release of constraint model.
Unfortunately, this basic pattern corresponds to a similar scenario that does not involve a release of constraint. It is commonly postulated that a trait’s evolutionary pattern may correspond best to a Brownian motion (BM) process without no central tendency imposed by the constraint in the OU model. If the basic Brownian rate parameter increases at the time of the innovation, a pattern of increased growth in disparity can still be observed without the corresponding mechanism of a release of constraint. Modeling a change in a Brownian rate parameter was first introduced in @O’Meara2006 in the software Brownie; hence we will refer to this as the Brownie model.
Though the processes are not identical, the exhibit remarkably similar patterns. Figure 2 illustrates the Brownie and release of constraint models through 500 replicate simulations of each. In the top panel, BM and OU processes with comparable parameters are shown for reference. In both the Brownie and release of constraint models, the same variance has been reached at the time of the shift and at the time the simulation ends. The distinguishing feature in the release of constraint model is similar to the distinction between a BM and OU processes – before the shift occurs, the trait dynamics have begun to approach an equilibrium that balances the diversification process against the constraint. This corresponds to traits values more closely reflecting a match to their environment than to their evolutionary history. After the shift in selection occurs, the traits begin to explore outside the range previously possible under the strong constraint.
By contrast, the Brownie model shows a sharper transition at this boundary. A close look at the figure shows a shock front at the moment of this transition.\ Whereas in the release of constraint model, the trajectories only loose their central bias, but otherwise continue to make similar step sizes, in the Brownie model the entire tempo of the evolutionary process has changed, taking bigger steps in both directions. It is these subtle differences in the patterns of the evolutionary processes implied by the different models that we seek to tease apart. To obtain the most powerful statistical comparison between the two models that accounts for the uncertainty in the model estimate, we use the method described in Boettiger, Coop, and Ralph (2012) which uses a bootstrap simulation approach to compare likelihood ratios of the models.
Results
Estimating the release of constraint model
Comparing models
(Show that Brownie model rejects the OUCH model/thetas?)
Discussion
Our analysis identified two functional traits in the labrid jaw morphology that show clear evidence of a release of constraint.
Functional innovations…
Acknowledgements
This work was supported by a Computational Sciences Graduate Fellowship from the Department of Energy under grant number DE-FG02-97ER25308 to CB and NSF grant DEB-1061981 to PCW.
References
Beaulieu, Jeremy M., Dwueng-Chwuan Jhwueng, Carl Boettiger, and Brian C. O’Meara. 2012. “Modeling Stabilizing Selection: Expanding the Ornstein-Uhlenbeck Model of Adaptive Evolution.” Evolution (mar). doi:10.1111/j.1558-5646.2012.01619.x. http://doi.wiley.com/10.1111/j.1558-5646.2012.01619.x.
Boettiger, Carl, Graham Coop, and Peter Ralph. 2012. “Is your phylogeny informative? Measuring the power of comparative methods.” Evolution (jan). doi:10.1111/j.1558-5646.2012.01574.x. http://doi.wiley.com/10.1111/j.1558-5646.2012.01574.x.
Butler, Marguerite A., and Aaron A. King. 2004. “Phylogenetic Comparative Analysis: A Modeling Approach for Adaptive Evolution.” The American Naturalist 164 (dec): 683–695. doi:10.1086/426002. http://www.jstor.org/stable/10.1086/426002.
Hansen, Thomas F., and E. P. Martins. 1996. “Translating between microevolutionary process and macroevolutionary patterns: the correlation structure of interspecific data.” Evolution 50: 1404–1417.
Price, Samantha a, Peter C. Wainwright, David R. Bellwood, Erem Kazancioglu, David C. Collar, and Thomas J. Near. 2010. “Functional innovations and morphological diversification in parrotfish.” Evolution; international journal of organic evolution 64 (oct): 3057–68. doi:10.1111/j.1558-5646.2010.01036.x. http://www.ncbi.nlm.nih.gov/pubmed/20497217.
R Package Documentation
Browse R Packages
We want your feedback!
Note that we can't provide technical support on individual packages. You should contact the package authors for that.
Embedding an R snippet on your website
Add the following code to your website.
For more information on customizing the embed code, read Embedding Snippets.